
|
| |||
|
Разделы
Главная
Сапромат
Моделирование
Взаимодействие
Методы
Инновации
Индукция
Исследования
Факторизация
Частоты
Популярное
Как составляется проект слаботочных сетей?
Как защитить объект?
Слаботочные системы в проекте «Умный дом»
Какой дом надежнее: каркасный или брусовой?
Как правильно создавать слаботочные системы?
Что такое энергоэффективные дома?
|
Главная » Квазитрехмерная модель 1 2 Квазитрехмерная модель динамики биомассы Галицкий В.В. (galvv@rambler.ru ) Институт физико-химических и биологических проблем почвоведения РАН Введение Понятие - свободно растущее растение - было использовано в работах (Галицкий, 1999; 2000; Galitskii, 2003) в качестве одного из основных элементов двумерных моделей растения и сообщества растений, предназначенных, в первую очередь, для изучения влияния свойств индивидуальных растений, составляющих сообщество, правил их взаимодействия и локальной структуры сообщества на его динамику или иными словами - проблемы микроскопической обусловленности макроскопического поведения сообщества. Свободно растущее растение по определению не испытывает влияния конкуренции за ресурсы со стороны других растений сообщества, но находится в экологических условиях, создаваемых сообществом. В упомянутых моделях оно является некоторым идеалом, относительно которого рассматривается поведение (угнетаемого) растения. Оно позволяет выделить и описать процесс собственно конкуренции из множества разнообразных процессов, в которых участвует и влияние которых испытывает растение в сообществе. Сходные понятия - отдельно , изолированно , открыто растущее растение - иногда рассматриваются в литературе (Сукачев, 1972; Dale et al., 1985). Отличие состоит в том, что там обсуждаются конкретные, существующие в природе объекты - растения, достаточно удаленные пространственно от других растений и потому не испытывающие конкуренции, но и находящиеся в условиях (в частности, микроклиматических), отличных, вообще говоря, от условий, имеющихся в сообществе взаимодействующих растений. Используемый же нами (умозрительный) объект - свободно растущее растение - находится в условиях сообщества, не испытывая конкуренции. По сути, характеристики свободно растущего растения могут быть использованы (теоретически, по крайней мере) в качестве такой меры нормальной производительности данных условий местопроизрастания или бонитета территории (Сукачев, 1972, с. 93), которая (мера) не была бы отягощена факторами, связанными с влиянием структуры и истории конкретного сообщества. В двумерном случае в качестве характеристик свободно растущего растения (его имитационной модели) были использованы две функции от возраста растения: Ap(T) - площадь, необходимая растению для свободного роста в возрасте T и Bp(T) - соответствующая биомасса (в Приложении А приводится краткое формальное описание 2D-модели). Под биомассой понимается физиологически активная часть полной массы растения (Галицкий, 1999; Galitskii, 2003). Форма площади в случае изотропной территории - круг увеличивающегося с возрастом радиуса. Получение этих функций, связывающих физиологию, морфологию и т.п. внутренние аспекты жизнедеятельности растительного организма с экологическими условиями его существования, исключая конкурентное взаимодействие с соседними растениями, должно представлять собой предмет других моделей и лежит вне рассматриваемой здесь задачи. Наша цель - использовать эти функции и для рассматриваемой далее квазитрехмерной модели. И. А. Полетаев (1966) с помощью несложной модели, основанной на вполне разумном предположении, показал, что рост дерева в высоту физически ограничен необходимостью подъема влаги и может быть описан как H(T) = H0 th(T/A1), (1) где A1 - параметр масштаба времени и th(x) - гиперболический тангенс. В соответствии с этим результатом в двумерных моделях (Галицкий, 1999; Galitskii, 2003) в качестве биомассы свободно растущего дерева было использовано BF (T) = B0 th1 (T/A1) (2) и AF (T) = A0 th2 (T/A1), (3) где H0, B0, A1;A0 - размерные параметры и \x - аллометрический параметр, в случае изометрии равный 2 при B-типе роста (Галицкий, 1999). Трехмерность для наших целей достаточно просто и наглядно (по крайней мере, для хвойных деревьев бореальных местообитаний) вводится путем выделения в биомассе дерева составляющих ее секций, каждая из которых может быть сопоставлена с суммарной биомассой одновозрастных ветвей, относящихся к одному узлу (с ветвями) ствола дерева, и соответствующего междоузлия. Такой морфологический способ выделения секций биомассы не единственный, но как будет видно, вполне конструктивный. Использование префикса квази в характеристике модели связано с тем, что в описываемой модели в действительности нет полноценного описания третьего измерения, т.к. рост дерева в высоту (как и периодичность появления секций) не определен внутри модели и не связан с ее функционированием, а задается снаружи, что впрочем на начальном этапе построения и анализа модели не существенно. В двумерной модели (Галицкий, 1999; 2000; Galitskii, 2003) биомасса является источником фитомассы - физиологически пассивной части массы растения. Динамика фитомассы описывалась простой моделью: биомасса с некоторой удельной интенсивностью тм 1 переходит в пассивное состояние, в фитомассу P dP/dT = B/тм . (4) При трехмерном рассмотрении в модели динамики фитомассы необходимо учитывать процессы транспорта ассимилятов (Курсанов, 1976) и соответствующая модель существенно сложнее. В работе (Галицкий, 2004) приведены некоторые результаты одного наивного варианта такого учета, качественно во всяком случае похожие на то, что наблюдается в действительности. Однако, это отдельный и сложный вопрос и здесь он не рассматривается, поскольку при заданной динамике биомассы свободно растущего дерева и при секционном представлении дерева динамику биомассы секций свободно растущего дерева и дерева в условиях ограниченности территории роста можно рассматривать без этого, расплачиваясь, в частности, добавлением в названии модели префикса квази. В работе (Галицкий, 2004) была описана квазитрехмерная секционная модель динамики биомассы свободно растущего дерева. Ниже кратко излагаются основные моменты этой модели, и затем приводится описание устройства основанной на ней квазитрехмерной секционной модели динамики биомассы дерева, растущего на некоторой ограниченной территории. Квазитрехмерная секционная модель динамики биомассы свободно растущего дерева В рамках общей концепции (Галицкий, Тюрюканов, 2001) движения от рассмотрения объекта под минимальным углом зрения к постепенной детализации представлений об его устройстве, вводя в рассмотрение новую деталь - секцию биомассы дерева, попытаемся установить ее соотношение с соответствующим объектом предыдущего уровня - биомассой всего дерева. Начиная с момента появления каждой новой i-ой секции (стволовые мутовка и междоузлие), дерево начинает реализовывать новую (которая может быть той же самой, что была у предыдущей секции) в определенной степени виртуальную динамику биомассы BFi(T-Ti) верхней части дерева, начинающейся с этой секции (BFri(x)=0, x<0). С нулевой секции начинает реализовываться BFr0(T) - виртуальная динамика биомассы всего дерева. Каждая из этих динамик действительна вплоть до появления новой следующей секции, а затем ее можно считать действительной в части, относящейся к собственно данной секции и виртуальной для следующей секции, поскольку следующая секция реализует аналогичным образом свою динамику. Очевидно, что биомасса всего дерева есть сумма биомасс всех его секций, имеющихся на данный момент времени. Предположим, что с момента появления на макушке дерева новой i-ой секции динамика части биомассы дерева, начинающейся с этой секции, описывается той же самой функцией свободного роста BF(T), что и для всех предыдущих секций дерева (в этом случае совпадающей с функцией для всего дерева), но сдвинутой на момент времени появления i-ой секции. Вплоть до появления следующей (/+1)-ой секции эта функция описывает также и динамику биомассы i-ой секции. То же самое можно сказать и о следующей (i+ 1)-ой секции, но уже с учетом сдвига на момент появления этой секции. После появления (i+ 1)-ой секции динамика биомассы i-ой секции может быть вычислена как разность биомассы дерева, начиная с этой секции, и биомассы дерева, начиная с +1)-ой секции. Таким образом, если секции биомассы дерева появляются с шагом по времени AT, то секция, появившаяся в момент Ti будет иметь зависимость биомассы от времени bF,(T) = BF,(T-T) - BFl+i(T-T,-AT), (5) где все функции от отрицательного аргумента равны нулю. Очевидно, что, просуммировав такие парциальные функции по всем имеющимся на данный момент секциям, мы получаем заданную функцию BF(T) для всего дерева. Из выражения (5) следует, что вид зависимости биомассы собственно секции от времени bFi(T) может качественно отличаться от зависимости BF(T) для всего дерева. Если полагать, что все дерево имеет -образную функцию BF(T) (B-тип роста, например, BF(T)= B0 th)i(T/A1), Галицкий, 1999), то секция будет иметь колоколообразную зависимость биомассы от времени, т. е. со временем биомасса секции (биомасса ветвей данного узла и соответствующего междоузлия) должна отмирать. Таким образом, нижние, более старые ветви дерева могут отмирать не только из-за конкуренции и других биологических обстоятельств, но и по математическим причинам. На рисунке 1 представлены зависимости от времени биомассы для нескольких номеров секций i=NL секций, рассчитанные с использованием выражений (2) и (5) при повторении секциями виртуальных зависимостей BFi(x) = BF(x), i=0,1, ..., x>0.  0,008 i = 0 50 100 \ \ 200 0,000 Рис. 1. Динамика биомассы некоторых секций свободно растущего дерева. Модель динамики биомассы дерева, растущего на ограниченной территории Как и в двумерном случае площадь доступной дереву территории будем считать мерой доступных дереву ресурсов. Рассмотрим азимутальную структуру биомассы дерева и свяжем ее с формой территории, на которой оно растет. Свободно растущее дерево очевидно имеет годограф равномерного азимутального распределения биомассы в форме круга увеличивающегося с возрастом радиуса. Вполне естественно предположить, что если по какому-то азимуту форма территории не обеспечивает выполнение условий свободного роста, то для этого азимута рост биомассы будет замедляться и, вообще говоря, соответствующая часть биомассы начнет отмирать. Натурные наблюдения над деревьями, растущими достаточно близко друг к другу, показывают, что нижние соприкасающиеся части крон между ними растут гораздо слабее, чем в противоположных направлениях, а рост верхних вообще говоря не зависит от азимута. 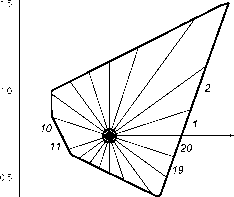 Рис. 2. Разбиение многоугольника (полигона) Вороного (Галицкий, Мироненко, 1981) на секторы, ns=20. Разобьем круг свободного роста (угол 2п) вокруг точки расположения дерева на ns одинаковых секторов (рис. 2) и к каждому у'-му сектору i-той секции применим уравнение, полученное при двумерном моделировании динамики биомассы дерева (Приложение А). Для этого разделим уравнение A3 на ns и заменим в линейных по B/ns и BF/ns выражениях B/ns и BF/ns на biJ- и bFij=bFi/ns - биомассау'-го сектора i-секции моделируемого дерева и биомасса у'-го сектора свободно растущей i-секции соответственно. В выражении А4 вместо площади роста A(T) будем использовать площадь a.ij(T-T) пересечения соответствующегоу'-го сектора круга свободного роста i-той секции и территории, доступной дереву, которая как и в двумерной модели сообщества (Галицкий, 2000; Galitskii, 2003) есть соответствующий полигон Вороного. Соответственно вместо AF(T) используем aFi(T)=AF(T-Ti)/ns . Множитель (B/BF)Y в выражении А6, не дающий биомассе дерева, бывшего в угнетении из-за недостатка площади для роста и внезапно освободившегося, расти как свободно росшее дерево того же возраста, следует заменить на (b /bFiy)r. Затраты на поддержание биомассы сектора определяются как произведение удельных затрат дерева, умноженных на биомассу сектора. В результате модель динамики биомассы у'-го сектора i-той секции дерева может быть записана в следующем виде dbt/dT=K(T-T)(btJ/r(B,T)+fi(btJ/bFij) dbFu/dT) - b4/r(B,T), (6) где K(T-T)=min(1, aij(T-T) / aF,(T)), (7) T-1(B,T)=(btJ cBk-1 + 1/TM) /(1+cg(1-k(T-T))), (8) /bF,i)=( btJ /bF ,)r / (1+cg(1-k(T-T))) , (9) B- биомасса всего дерева, Ti- момент появления i-той секции (Ti+1=Ti+AT), i=0,1,...,у'=1,..., ns. Аналогично условию отмирания дерева в двумерной модели (Галицкий, 1999) сектор и секция считаются отмершими, когда их биомассы удовлетворяет соответственно неравенствам by < su bp , b, < su bF . (10) Биомасса секции дерева есть сумма биомасс составляющих секторов bi 1- b, (11) и биомасса дерева есть сумма биомасс имеющихся на данный момент секций B= I. Ь, . (12) Примеры квазитрехмерной динамики биомассы дерева Расчеты проводились для дерева, растущего на полигоне, изображенном на рис. 2. Параметры модели дерева были взяты следующие: A0=100.0, A1=100.0, B0=1.0, =2.37, <c=0.01, k=0.8, тм=0.1, cg=0.1, y=0.24, AT=1.0, число секторов ns=20. Далее в обозначениях биомасс сектора bij и секции bi индексы есть соответственно i -номер секции и у - номер сектора. На рис. 3 приведены расчетная динамика для сектора 3 секции 0 для нескольких значений масштабного множителя ksc для линейных размеров полигона, где ksc=1 соответствует размерам полигона, изображенного на рис. 2. Огибающая динамик секторов представляет динамику деленной на число секторов ns биомассы свободно растущей секции. На рис. 4 приведена динамика биомассы секции 0 дерева для нескольких значений масштабного множителя ksc . На рис. 5 представлены динамики биомассы секторов 3 и 11, существенно отличающихся по величине площади, при ksc=1. Одним из результатов q3D-модели являются годографы азимутального распределения биомассы секции. На рис. 6 изображены несколько таких 4,0x10 2,0x10-4 0,0-
0,006 0,004 0,002 0,000 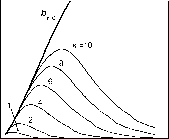 0 20 40 60 80 100 Рис. 3. Динамика биомассы сектора 3 (рис.2) Рис. 4. Динамика биомассы секции 0 секции 0 при различных значениях ksc. при различных значениях ksc. 4,0x1 0-: 2,0x1 0-:
Рис. 5. Динамика биомассы секторов 3 и 11 секции 0 (см. рис. 2). о 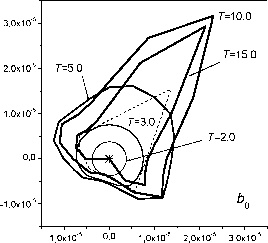 Рис. 6. Годографы распределения биомассы 0-секции для ряда моментов времени. Штриховой линией показан полигон Вороного. годографов 0-секции дерева для ряда моментов времени. Можно видеть, что годограф азимутальной плотности распределения биомассы секции, имея вначале круговую форму, со временем деформируется в соответствии с расстоянием от дерева до границы полигона в соответствующем направлении. Для рассматриваемой здесь упрощенной модели последующие секции дерева повторяют эту картину с соответствующим сдвигом по времени. На рис. 7 представлены в виде kite-диаграмм распределения биомассы по секциям i для различных возрастов дерева. Рисунок иллюстрирует движение волны биомассы (кроны) дерева в высоту путем нарастания новых секций на макушке дерева и отмирания старых снизу кроны. Необходимо напомнить, что азимутальное распределение биомассы для нескольких секций изображено на рис. 6.
4,0x10 4 2,0x10 4 0,0 2,0x10 4 4,0x10 b. i Рис. 7. Динамика распределения по секциям i биомассы дерева, растущего на полигоне (рис. 2, ks=1), для ряда моментов времени. 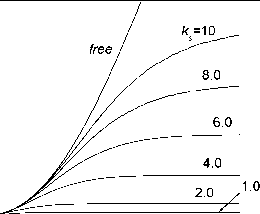 0 20 40 60 80 100 Рис. 8. Динамика биомассы дерева, растущего на полигоне (рис. 2) при различных значениях ksc. На рис. 8 приведена динамика биомассы дерева, занимающего изображенный на рис. 2 полигон, для различных значений линейного масштабного коэффициента ksc . Видно, что с увеличением линейных размеров полигона (увеличением его площади) растет величина стационарной биомассы дерева. В случае двумерной модели дерева (Галицкий, 1999) недостаток площади приводит к непосредственной и неминуемой гибели модельного дерева. В отличие от двумерной модели в трехмерном случае при недостаточной площади полигона, на котором растет дерево, отмирают нижние секции его биомассы, но прирастают верхние, вновь появляющиеся. Если высота дерева не играет никакой роли в его росте, то биомасса дерева выходит на стационарный уровень и дерево не отмирает при любом малом полигоне. 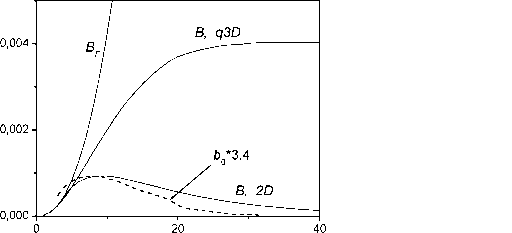 Рис. 9. Зависимости от времени биомассы для q3D- и 2D-моделей дерева, растущего на полигоне (рис. 2), 0-секции q3D-модели и для свободно растущего дерева. Представляет интерес сравнить поведение трехмерной (q3D) модели и ее прототипа - двумерной (2D) модели без учета формы полигона. На рис. 9 представлены динамики биомассы дерева для этих моделей, а также для свободно растущего дерева и для 0-секции q3D-модели при одинаковых общих параметрах моделей. Поведение динамик для модельных деревьев в обоих случаях различно -как отмечалось выше, при недостаточной площади для роста 2D модель непременно приводит к гибели модельного дерева, q3D модельное дерево никогда не отмирает. В то же время динамика для 2D-модели дерева довольно близка к динамике для 0-секции q3D-модели. Заключение Разработка трехмерных моделей дерева и сообщества деревьев, выполняемая в рамках принципа минимального угла зрения (Галицкий, Тюрюканов, 2001), представляет собой попытку продвинуться в понимании и описании механизма конкуренции в сообществе деревьев и использовать модели для рассмотрения эффектов и ситуаций, для которых двумерные модели оказались недостаточными. Двумерные модели (с учетом формы территории, на которой растет дерево, и без учета (Галицкий, 1999; 2000; Galitskii, 2003)) в принципе давали вполне адекватные (по крайней мере на качественном уровне) описания рассматривавшихся с их помощью ситуаций, хотя в некоторых областях пространства параметров моделей проявлялись заметные количественные расхождения с данными натурных наблюдений (Галицкий, 1998; 2003). Естественная попытка детализировать двумерную модель путем учета азимутального распределения биомассы двумерных деревьев в сообществе приводит к качественно неадекватному результату - при смыкании крон кроновый полог модельного сообщества становится дырявым (Галицкий, не опубликовано), что очевидно не соответствует реальности для деревьев, но весьма похоже на ситуацию с нижними частями крон достаточно близко расположенных друг к другу соседних деревьев. Это показывает, что верхние части конкурирующих деревьев растут свободно, а нижние собственно и участвуют в конкуренции, т.е. при более детальном рассмотрении конкуренции между деревьями их следует рассматривать как трехмерный объект. Систематическое использование этого наблюдения приводит к представлению о необходимости выделения секций в морфологической структуре дерева как элемента модели. Как и в двумерной модели в представленной квазитрехмерной модели эффективно используется представление о свободно растущем дереве, позволяющее вычленить из множества процессов, структур и характеристик, влияющих на рост дерева те, которые непосредственно связаны с конкуренцией с соседями в сообществе, а все остальное учесть в свойствах свободно растущего дерева. Секционная модель динамики биомассы свободно растущего дерева основывается на двух исходных положениях - iS-образная форма динамики биомассы дерева и очевидная процедура периодического появления на макушке дерева виртуальных деревьев. В результате получен наглядный эффект оголения снизу ствола дерева от ветвей. Насколько нам известно, iS-образность динамики биомассы дерева в натурных исследованиях древостоев непосредственно не фиксировалась, что в первую очередь может быть связано с чрезмерной длительностью необходимых для этого наблюдений. Таблицы хода роста дают выпуклую вверх кривую динамики запаса древесины (фитомассы). Полученный с помощью секционной модели результат - оголение снизу ствола свободно растущего дерева может рассматриваться при достаточной длительности наблюдения в качестве необходимого признака iS-образности динамики биомассы для конкретных видов деревьев. Квазитрехмерная секционная модель динамики биомассы дерева, растущего не свободно, т. е. растущего на ограниченной территории, пересечение которой с кругом свободного роста, начиная с некоторого возраста, не совпадает с этим кругом, сконструирована с использованием техники, примененной при разработке двумерной модели, учитывающей форму полигона (Galitskii, 2003) и азимутальную структуру биомассы дерева. Представленная здесь квазитрехмерная модель динамики биомассы дерева очевидно не является более или менее законченной моделью динамики биомассы дерева и тем более дерева в целом. Это видно, например, из рисунка 8 - q3D дерево не отмирает ни при каком угнетении (недостатке площади для роста). Однако, это обстоятельство, как и описанная выше т.н. дырявость кронового полога в двумерной модели, является причиной и поводом к дальнейшему поиску пропорционально уравновешенной трехмерной модели. Это будет сделано в последующих публикациях. Здесь можно отметить, что желательность пропорциональности разных сторон модели может рассматриваться как еще один наряду с упомянутым принципом минимального угла зрения существенный принцип моделирования. Работа выполнена при поддержке Российского фонда фундаментальных исследований, грант №03-04-48616. Литература 1. Галицкий В. В. 1984. О влиянии отчуждения части фитомассы на рост растения Известия АН. Сер. биол. № 6, 823-833. 2. Галицкий В.В. 1999. Моделирование сообщества растений: индивидуально- ориентированный подход. I. Модель растения Известия АН. Сер. биол. № 5. 539-546. 3. Галицкий В. В. 2000. Моделирование сообщества растений: индивидуально- ориентированный подход. II. Модель сообщества Известия АН. Сер. биол. № 2. 178-185. 4. Галицкий В.В. 2003. Динамика конкуренции в растительных сообществах различной степени однородности. Электронный журнал Исследовано в России , 2207-2221, http: zhurnal.ape.relarn.ru/articles/2003/184.pdf. 184 5. Галицкий В. В. 2004. Квазитрехмерная модель свободно растущего дерева Электронный журнал ИССЛЕДОВАНО В РОССИИ . 247, 2646-3662. http: zhurnal.ape.relarn.ru/articles/2004/247.pdf 6. Галицкий В.В., Мироненко Е.В. 1981. Мозаика Вороного на плоскости. Алгоритм построения. Пущино. ОНТИ НЦБИ АН СССР. 7. Галицкий В.В., Тюрюканов А.Н. 2001. О методологических предпосылках моделирования в биогеоценологии В кн.: А.Н. Тюрюканов. Избранные труды. М.: Изд-во РЭФИА. 94-108. 8. Курсанов А.Л. 1976. Транспорт ассимилятов в растении. М. Наука. 9. Полетаев И. А. 1966. О математических моделях биогеоценотических процессов. В: Проблемы кибернетики. Вып. 16. М.: Наука. 175-177. 10. Сукачев В.Н. 1972. Избранные труды. Т. I. М.: Наука. 11. Dale V.H., Doyle T.W., Shugart H.H. 1985. A comparison of tree growth models Ecol. Modelling. 29: 1145-169. 12. Galitskii V.V. 2003. The 2D modeling of tree community: from microscopic description to macroscopic behavior For. Ecol. & Manag. 183. Iss. 1-3. 95-111. 13. Makela, A., Hari, P. 1986. Stand growth model based on carbon uptake and allocation in individual trees. Ecol. Model. 33: 315:331. 1 2 |
|
| |||